
RESEARCH 植物の生活史の進化
(張列弛 院生、Ruan de Villiers 院生、青山剛士 研究員、石川雅樹 助教他:大学院生募集中)
ヒトは生涯を通じて基本的体制が変化しません。しかし、陸上植物は生活史の中で体制を変化させます。例えば、コケ植物のヒメツリガネゴケは糸状の原糸体、茎葉を作る茎葉体、棍棒状の胞子体と生涯にいろいろな形の体を作ります。このような体制の転換は、どのような仕組みで制御され、進化してきたのでしょうか。
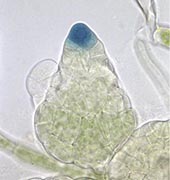
ヒメツリガネゴケの原糸体でポリコーム抑制複合体2 遺伝子を壊すと、通常は、受精したあとに形成される胞子体が、受精無しで形成される Okano et al. 2009。
(詳しい研究内容)
動物と植物の大きな違いは、植物には1倍体と2倍体世代があり、両者が交代する生活史を持つことです。受精卵から2倍体の発生プログラムに従って2倍体の体が形成され、減数分裂によってできた胞子から1倍体の発生プログラムが開始して1倍体の体ができます。ところが、1倍体と2倍体の発生プログラムは受精や減数分裂と関係無く誘導できることが知られていました。例えば、ベニシダなどのシダ類は、1倍体である前葉体の細胞が受精せずに2倍体の発生プログラムを開始して2倍体である胞子体を形成することが知られており、アポガミー(無配生殖)と呼ばれています。また、コケの中には2倍体である胞子体から減数分裂無しに1倍体発生プログラムが開始し、1倍体である原糸体が生じる現象が知られており、アポスポリー(無胞子生殖)と呼ばれています。これらのことは、1倍体と2倍体の発生プログラムを開始する何らかのスイッチがあることを示唆していますが、その実体は不明でした。ところが数年前、ヒメツリガネゴケで別な目的のため、クロマチン修飾を制御するポリコーム抑制複合体2遺伝子を欠失させたところ、なんと、1倍体である原糸体から受精無しで2倍体である胞子体が生えてきてしまったのです。これが、ポリコーム抑制複合体2が2倍体発生プログラムの開始を制御していることの発見でした(Okano et al. 2009)。
Okano, Y., Aono, N., Hiwatashi, Y., Murata, T., Nishiyama, T., Ishikawa, T., Kubo, M., and Hasebe, M. (2009). A polycomb repressive complex 2 gene regulates apogamy and gives evolutionary insights into early land plant evolution. Proc. Natl. Acad. Sci. USA 106: 16321-16326.
榊原恵子 博士(現・金沢大学)は葉の形態進化を研究したいと言って私の研究室に大学院生としてやってきました。葉の形態進化に関わる遺伝子はまだ良く分かっておらず、とりあえず、ホメオボックス遺伝子の機能解析を行いました。しかし、葉の形態ではなく、予想外に仮根形成を制御する因子だということがわかり、仮根形成の初めての分子機構解明ということで、学位を取得しました(Sakakibara et al. 2003. Development 130: 4835-4846)。榊原 博士はその後もホメオボックス遺伝子の研究を、学振特別研究員としてオーストラリアの Bowman 教授研究室、ERATO 分化全能性進化プロジェクト技術参事として基生研、そして、出口博則 教授研究室助教として広島大で続けました。そして、これまで機能が全く未知だった class 2 KNOX ホメオボックス遺伝子をヒメツリガネゴケで欠失させたところ、成熟した胞子体ができないことに気づき、さらに、その未熟な胞子体を詳細に観察したところ、なんと、減数分裂によって胞子からできるはずの原糸体が、減数分裂無しで胞子体上から生えてきていることを見つけました。すなわち、class 2 KNOX ホメオボックス遺伝子が1倍体発生プログラムの開始を制御していることがわかりました(Sakakibara et al. 2013)。
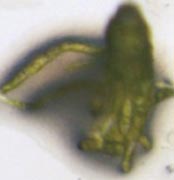
ヒメツリガネゴケ胞子体でclass 2 KNOXホメオボックス遺伝子を壊すと、本来は減数分裂後の胞子から形成される原糸体が、減数分裂無しで形成される(Sakakibara et al. 2013 Science)。
Sakakibara, K., Ando, S., Yip, H.K., Tamada, Y., Hiwatashi, Y., Murata, T., Deguchi, H., Hasebe, M., and Bowman, J.L. (2013). KNOX2 genes regulate the haploid-to-diploid morphological transition in land plants. Science 339: 1067-1070.
ヒメツリガネゴケの1倍体世代は胞子細胞が分裂して原糸体幹細胞を形成し、この幹細胞が1倍体世代の源になります。一方、2倍体世代は受精卵が分裂して胞子体幹細胞を形成し、この幹細胞が2倍体世代の源となります。つまり、世代交代とは異なった性質の幹細胞を作ることにあります。さらに、1倍体世代において、原糸体幹細胞からできた原糸体細胞から茎葉体幹細胞が形成され茎葉体ができます。茎葉体幹細胞は低温条件で造精器幹細胞と造卵器幹細胞へと変化し、造精器と造卵器が形成されます。これらのことから、植物の生活史とは次々に異なった性質の幹細胞を適切な場所と時間に作り出すことなのだということに気づきました(Kofuji and Hasebe 2014)。すなわち、生活史がどのような分子機構によって制御されているか、また、どのような変化によって進化したかを知るには、異なった幹細胞を作り出す分子機構を調べることが解決の道だということになります。
Kofuji, R., and Hasebe, M. (2014). Eight types of stem cells in the life cycle of the moss Physcomitrella patens. Curr. Opin. Plant Biol. 17: 13-21.
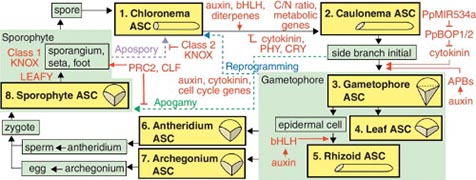
ヒメツリガネゴケの生活史。異なった種類の幹細胞が適当な場所と時間にできることで生活史が進行する。
総研大大学院生だった青山剛士 博士(現当研究室 研究員)は大学院に入ってくるときに「配偶体世代にできるコケの茎葉が胞子体世代にできる被子植物の茎葉とどう違うのかを調べたい」という明確な目標を持っていました。いくつかの遺伝子を調べたり、突然変異体を作ってみたりしていましたが、なかなかうまくいきませんでした。そんなとき、PLETHORA/AP2/BABYBOOM (APB)3重遺伝子欠失体は表現型がはっきりしなかったのですが、ゲノム解読の結果、APBにもう一つパラログがあることがわかり、4重遺伝子欠失体を作ったら、なんと茎葉体が出来なくなってしまいました。APB が茎葉体幹細胞形成のマスターレギュレーター、もう少し詳しく書くと、オーキシンとサイトカイニンと協調することで原糸体幹細胞と茎葉体幹細胞の作り分けを行う因子であることがわかったのです(Aoyama et al. 2013)。「知りたい」と強く思って実験していると、不思議と青い鳥がいて、かなってしまうことが多いような気がします。APB を中心とした遺伝子ネットワークをシダ植物や種子植物と比較することで、陸上植物の生活史、即ち、幹細胞転換機構の進化過程が推定できるはずだと考えています。
Aoyama, T., Hiwatashi, Y., Shigyo, M., Kofuji, R., Kubo, M., Ito, M., and Hasebe, M. (2012). AP2-type transcription factors determine stem cell identity in the moss Physcomitrella patens. Development 139: 3120-3129.
ヒメツリガネゴケの原糸体幹細胞は次々と分裂をして原糸体細胞を創り出します。原糸体細胞は原糸体幹細胞から2細胞ほど離れると幹細胞化がおこり、原糸体幹細胞と茎葉体幹細胞のどちらかを作ります。ここで茎葉体幹細胞になるスイッチを入れている遺伝子が先述の APB 遺伝子です。原糸体幹細胞は緑藻類に似て前期前微小管束形成を伴わない細胞分裂様式を持ち糸状の組織を作ります。一方、茎葉体幹細胞は前期前微小管束形成を伴う維管束植物型の細胞分裂を行い、シュートを形成します。従って、この幹細胞の性質決定機構は、幹細胞制御という点に加えて、陸上植物の細胞分裂機構と体制の進化解明の鍵となりそうです。さて、両者はどのように作り分けられるのでしょうか。これまでの予備的実験から、APB 遺伝子の発現は、オーキシン極性輸送体の分布制御によって引き起こされているらしいことがわかってきました。また、オーキシンとは別な植物ホルモンであるストリゴラクトンによっても制御されている可能性が示唆されています。南アフリカからの国費留学生の Ruan de Villiers 君がこの問題に取り組んでいます。